Maduración, activación y diferenciación de los Linfocitos
T
Grupo #4
El receptor clonotipico de las T (TCR) presenta dos
funciones principales segun la fase de desarrollo en que se encuentre la célula
dentro del linaje de los linfocitos T:
1.
Durante
la maduración de los timocitos en el timo, participa en la sleccion timica
positiva y negativa.
2.
Una
vez que el linfocito T ha madurado, emigra a la periferia, y entonces el
receptor participa en el reconocimiento de antígenos, lo que desencadena un
programa de activación que lleva a la proliferación y diferenciación de las
células T en dos subciones: uno de células efectores, y otro de células de
memoria.
El proceso de
reconocimiento varía según hablemos de TCR-2 (α β) o del TCR (ʏ δ): en el primer caso, y como ya hemos visto, se
reconocen peptides en el contexto del haplotipo propio del MHC clasico, y se
requieren moléculas coestimuladoras y coseñlizadoras, notablemente la CD4 (para
linfocitos Tн) o la CD8 (para los linfocitos Tc); en el caso del ʏ δ, no se require MHC clasico, y no participan CD4
ni CD8.
Los linfocitos con receptores de tipo α β, se dan 2 procesos 1 de
maduracion y otro de activación:
·
Maduracion:
la enorme diversidad antigénica potenciual se reduce a un 2% durante la
maduración intratimica de los timocitos: solo llegan a madurar aquellas células
restringidas a reconocer lo no-propio en el contexto del haplotipo MHC propio
(autorrestriccion y autotolerancia).
·
Activacion:
la activación de células T maduras periféricas se inicia con la inteccion entre
el TCR y un péptido antigénico enclavado en la hendidura del MHC. La baja
afinidad (10⁻⁵M) de esta interaccion ternaria se ve potenciada por la
presencia de correceptores y otras moléculas de membrana, que funcionan para
fortalecer la interracion ternaria TCR-peptido-MHC, y para transducir la señal
activadora al interior de la célula T.
Maduracion de las células T
Desarrollo del timo:
El estroma
timico surge al inicio del desarrollo embrionario a partir de capas
ectodérmicas y endodérmicas procedentes del tecer bolsillo faríngeo y de la
tercera hendidura branquial. Estas dos
estructuras se invaginan, y se cierran, y las dos capas quedan superpuestas, de
modo que la ectodérmica rodea a la endodérmica, formando el llamado rudimento
timico.
La capa
ectodérmica formara los tejidos epiteliales corticales del timo, mientras que
la capa endodérmica formara los tejidos epiteliales medulares.
En la corteza
encontramois solo timocitos en fases tempranas de su maduración, junto con
algún macrófagos, dentro del estroma cortical a base de células corticales
epiteliales. En la medula encontramos timocitos en fases mas acanzadas de
maduración con células dendríticas macrófagos, todos inmersos en un estroma
medular a base de células epiteliales medulares.
Rutas de desarrollo en el linaje de las células T
El primer
marcador de superficie en aparecer es el Thy-1 (equivalente al CD2 de humanos),
que ya no se pierde (por lo tanto, se trata de un marcador que caracteriza al
linaje de T). Estas células Thy-1+ CD4⁻ CD8⁻ (dobles negativas) pueden escoger dos vías alternativas:
1. En una de las dos rutas, las células hacen reordenaciones
productivas de ʏ y δ y expresan CD3 en su membrana. Suponen solo <1%
de los timocitos. Son las primeras en aparecer: se detectan al día 14 de
gestación, pero desaparecen al nacimiento.
2. La mayoría escoge una vía
alternativa, que discurre de la siguiente manera:
·
Día
16°: las células reordenan genes de cadenas β. Si no se logran reordenaciones productivas, entran en
apoptosis. Si la reordenación es productiva, la cadena β se asocial con la llamada cadena α sustitutiva, generando el receptor pTα: β (junto con el CD3). Este receptor induce la
proliferación celular y la coexpresion de CD4 y CD8: de este modo aparecen los
timocitos grandes doble positivo.
·
Día
17°: CD4+ CD8+ TCR-2+ (α β) CD3+. Se trata de los pequeños timocitos dobles
positivos, que dejan de dividirse. Estas células ya provistas del complejo
receptor especifico van a ser sometidas, hasta la época del nacimiento, a
selección positiva y negativa:
·
Selección positiva: sobreviven aquellas células
que tengan TCR capaces de reconocer MHC-I o MHC-II de células epiteliales del
timo.
·
Selección negativa: de aquellas células que han
pasado la selección positiva mueren por apoptosis las que posean TCR que
reconozcan con alta afinidad péptidos propios enclavados en el MHC o MHC propio
solo.
Los timocitos dobles positivos que superan la doble selección timica se desarrollan
en una de dos posibles alternativas:
·
CD4+
CD8⁻ TCR-2+ CD3+ (representan el; 10% de timocitos)
·
CD8+
CD4⁻ TCR-2+ CD3+ (un 5% de los timocitos)
·
Adicionalmente,
y quizás procedente de los anteriores, al
5° día del nacimiento se detecta una tercera subpoblacion de CD4⁻ CD8⁻ TCR+ CD3+
Localización intratimica de las diversas fases madurativas:
·
Los
timocitos doble negativos se localizan en la zona subcapsular de la corteza.
·
Pequeños
timocitos dobles positivos se localizan en la corteza.
·
Los
timocitos maduros CD4+ y CD8+ se ubican en la medula.
·
En
la corteza, las células epiteliales corticales establecen contactos por sus
largos procesos de membrana con los timocitos.
Selección timica positiva y
negativa
En ambos procesos selectivos parecen jugar un papel importante las células
de; estroma timico: células epiteliales timicas, macrófagos y células
dendríticas; todas ellas expresan en sus membranas grandes niveles de moléculas
MHC-I y/o MHC-II. Los timocitos inmaduros dobles positivos (CD4+ CD8+ TCR+
CD3+) interaccionan, por mecanismos aun oscuros, con estas células estromales,
lo que conducen a la selección positiva y negativa.
En la selección positiva se da interacción de los timocitos con células
epiteliales corticales del timo, algunos autores han sugerido la interacción de
los timocitos inmaduros dobles positivos con dichas células epiteliales por
medio del TCR restringido por MHC podría conllevar algún tipo de señal
protectora que librara a estos timocitos de la muerte celular programada.
En la selección negativa ocurren en la zona de transición cortico-medular y
en la medula timica, y en la que las células dendríticas y los macrófagos
interaccionan con los timocitos portadores de TCR de alta afinidad hacia (autopéptidos-MHC)
o hacia MHC solo.
La autotolerancia se consigue eliminando células T autorreactivas, y
permitiendo el desarrollo de las específicas que reconocen péptidos extraños
enclavados en el MHC propio.
Activación de los linfocitos T
coadyuvantes
La activación y expansión clonal de Tн es un acontecimiento central en la
producción de las respuestas inmunes especificas. Se trata de un proceso
complejo que en los últimos años está siendo paulatinamente desentrañando.
Los linfocitos T vírgenes son células en reposo que se encuentran
“aparcadas” en la fase G 0 del ciclo
celular. La activación, proliferación y diferenciación de estas células es un
fenómeno complejo.
La activación se inicia cuando el linfocito Tн interacciona, a través de su
complejo TCR-CD3, con el antígeno peptidico (exógeno) –procedente de
procesamiento endosomico- enclavado en el surco de MHC-II de una célula
presentadora.
Esta interacción inicial “dispara” una compleja cascada de acontecimientos
bioquímicos, en la que son genes, entre los que se cuentan el de la Il-2 y el
de su receptor.
La secreción autocrina de IL-2 por parte de los linfocitos Tн hace que
estos salgan de la fase G 0 y entren y progresen en el ciclo celular: ello
provoca la proliferación y diferenciación de la célula T en dos subpoblaciones:
una de células efectores (las T coadyuvantes o colaboradoras) y las Tн de
memoria.
Para que ocurra esto se requieren, además señales coestimulatorias. Si
tales señales químicas no se suministran al tiempo en que se está produciendo
la interacción especifica TCR- péptido –MHC, se induce un estado de incapacidad
de respuesta inmune que se denomina anergia, que se manifiesta en tolerancia
inmunológica hacia el estimulo antigénico.
Rutas de señalización intracelular
El TCR tiene colas citoplasmicas cortas que por sí mismas son incapaces de
señalización intracelular. Una vez que dicho TCR se une al péptido: MHC, esta
señal se transduce al interior de la célula T por medio de los dominios
citoplasmicos de CD3, el correceptor CD4 y varias moléculas accesorias (CD2,
CD45). Dicha transducción de señal se realiza por medio de una serie
protein-quinasas y protein-fosfatasa.
Algunas enzimas de la ruta de
señalización.
Protein-quinasas de la familia
del protooncogen src.
1. Proteína
p56 ıcκ:
·
se
trata de una protein-quinasa que se une a membrana mediante acido miristico
engarzado a la glicola en posición 2 (Gly2).
·
Posee
dos secuencias homólogos con otras proteínas (SH2 y SH3).
·
La
porción carboxiterminal es la que tiene actividad de quinasa. Obsérvese la
existencia de dos tirosinas (representadas por Y); la que está en la posición
394 (denominada de regulación positiva) es la tirosina que se fosforila al
activarse el linfocito T, mientras que la que está en posición 505 (llamada Tyr
de regulación negativa) esta fosforilada (Tyr-P) células T en reposo, y se
desfosforila cuando las células se activan.
·
En
el primer tercio se encuentra una cisteína que será la encargada de unirse por
puente disulfuro con CD4 (o en el caso Tc con CD8). También se asocia físicamente
con las cadenas ξ y ε del CD3.
2. Proteína
p59 fyn:
·
Su
estructura es muy parecida a la de p56 ick. También se encuentra anclada a la
membrana por miristilacion.
·
Igualmente
posee una Tyr cerca del extremo carboxil-terminal, que cuando esta fosforilada
hace que la p59 fyn esta inactiva, y otro sitio Tyr capaz de recibir fosfato
por autofosforilacion de esta quinasa, lo cual hace que la proteína pueda
fosforilar a otras proteínas.
·
Esta
físicamente asociada a cadenas ξ del CD3.
3. Fosforilasa
ZAP-70
·
No
está asociada por miristilacion a la membrana.
·
Contiene
una Tyr capaz de autofosforilarse, pero a diferencia de las proteínas de la
familia src, carece Tyr de regulación negativa.
·
En
las células T en reposo, la ZAP-70 no se encuentra asociada al complejo TCR-CD3;
sin embrago, cuando se inicia el proceso de activación celular, y una vez que
las cadenas ξ y ε de CD3 quedan fosforiladas por otras protein-quinasas,
a ZAP-70, por medio de sus dominios SH2 se une a estas cadenzas fosforiladas, y
entonces queda activada en su capacidad de fosforilasa.
4. Fosfatasa
CD45 (=LCA=T200)
·
El
CD45 es en realidad una familia de fosfatasas especificas de tirosina, que
aparecen en todas las células del linaje hematopoyético excepto en los
eritrocitos.
·
Existen
varias isoformas, de entre 180-200 kDa, que proceden de procesamiento
alternativo de un mismo tipo de ARN, y cada una de ellas aparece en
determinados tipos celulares.
·
Tiene
un dominio extracelular, que esta glucosilado; se une a la CD22
·
Su
porción citoplasmica es larga, y cuenta con dos dominios dotados de activad
fosfatasa de tirosinas (PTP).
·
Parece
ser que una de sus funciones es desfosforilar la Tyr-p situada cerca del
extremo carboxi-terminal de las protein-quinasas (PTK) p56ick y p59fyn.
5. Modelo
actual de la activación del linfocito Tн
·
La
señalización a través del complejo TCR-CD3 requiere que se agreguen muchos
complejos junto con sus correspondientes, correceptores CD4, y con CD45. Los
numerosos conjuntos TCR-CD3-CD4 interaccionan simultáneamente con muchos
complejos péptido: MHC-II de la célula presentadora de Ag.
·
La
actividad fosfatasa de CD45 provoca la desfosforilacion de la tirosina
fosforilada (Tyr-P) carboxil-terminal de p56ick y de p59fyn, lo que se supone
la activación de estas dos protein-tirosinquinasas (PTK): se autofosforilan en
la otra tirosina.
·
La
activación de las dos PTK citadas por autofosforilacion provoca que a su vez
estas fosforilen las cadenas del complejo CD3, reconociendo las secuencias ARAM
en ξ y en ε. También se fosforila la cola a las colas fosforiladas
de CD3 y CD4 se une ahora la ZAP-70, de modo que esta adquiere a su vez su
actividad de proteinquinasa, con lo que puede fosforilar a cadenas del CD3 y a
otras proteínas.
·
La ZAP-70 activa y la Fyn activa fosforilan a la
fosfolipasa Cʏ1 (PLCʏ1), que originalmente es una proteína citoplasmica; al
fosforilarse la PLCʏ1 se activa y emigra al lado citoplasmico de la membrana,
reconociendo otras proteínas que tienen tirosinas fosforiladas.
·
La PLCʏ 1 hidroliza a este PIP2, generando
inositol-trifosfato (IP3) y diacilglicerol (DAG), cada uno de los cuales
suponen el arranque de sendas rutas dentro de esta compleja cascada activadora:
a)
Ruta del inositol-trifosfato (IP3):
§ El Ip3 se une a un receptor especifico situado en el REr,
provocando la salida al citoplasma de grandes cantidades de Ca++, y junto con
IP4 provoca también la entrada desde el exterior celular, a través de canales
de calcio de la membrana citoplasmica, de mas cantidades de este catión.
§ El aumento intracelular Ca++ estimula a la enzima
calmodulina, que es una serin/treonin-quinasa.
§ La calmodulina activada activa a su vez a la
calcineurina, que es una fosfatasa.
§ La calcineurina activada cataliza la desfosforilacion del
factor NF-AT citoplasmico fosforilado (NF-ATc-P).
§ Una vez desfosforilado, el NF-AT emigra al núcleo, donde
se junta con el factor nuclear AP1, formando entrambos un factor de activación
transcripcional de varios genes, entre ellos el que codifica la citoquina IL-2.
b)
Ruta del diacilglicerol (DAG):
§ El DAG estimula, junto con el Ca++, a la protein-quinasa
C (PKC), que hasta ese momento residía en el citoplasma.
§ Al activarse, la PKC emigra a la cara interna de la
membrana citoplasmica; allí, en presencia de los fosfolipidos, ejerce su
función como serin/treonin-quinasa:
·
Fosforila
una amplia variedad de proteínas, entre las cuales se encuentra la codificada
por el protooncogen ras. La proteína Ras a su vez inicia otra cascada de
fosforilaciones que llega hasta l quinasa <AP.
·
Otra
de las consecuencias de la actividad PKC es que se fosforila el componente
inhibidor del factor kB que estaba retenido en el citoplasma. Al fosforilarse,
el componente inhibido queda a merced unas proteasas, que lo degradan. Es entonces
cuando el NF-kB puede emigrar al núcleo y unirse secuencias específicas del promotor
del gen IL-2 y de otros genes.
La señal coestimulatoria
Además de las señales suministradas a partir del contacto entre el complejo
TCR-CD3 con el péptido-MHC, la activación del linfocito Tн requiere una señal
adicional, denominada coestimulatoria, que puede consistir en alguna de las
siguientes:
·
La
citoquina IL-1, suministrada por la célula presentadora de antígeno (APC).
·
La
citoquina IL-6, de la APC.
·
Pero
la señal más potente es la que supone el contacto entre la molécula B7 (=CD80)
de la célula presentadora y la CD28 o la CTLA-4 del linfocito Tн.
B7 (=CD80)
consta de dos cadenas idénticas con dos dominios de tipo Ig. Se expresa
exclusivamente en células presentadoras de antígeno capaces de estimular a
linfocitos T.
La CD28 es una
glucoproteina homodimerica, cuyo monómero pesa 44 KDa, presente en linfocitos
Tн.
La CTLA-4 esta
codificada por un gen cercano al de la CD28, presentando ambas grandes
homologías. Pero la CTLA-4 solo se expresa en linfocitos Tн activados, siendo
su afinidad muy alta hacia la molécula B7.
Activación génica
Los genes se
activan y se pueden clasificar según el momento relativo de su expresión, en 3
categorías:
A. Genes de expresión inmediata (media hora). Estrictamente
hablando, estos genes no se activan, sino sus productos ya preformados.
B. Genes de expresión temprana (1 a 2 horas): son
esencialmente los que codifican las citoquinas IL-2 (así como el gen de su
receptor IL-2R), IL-#, IL-6 e interferon gamma (IFN⁻ʏ).
C. Genes de expresión tardía (hasta 2 días o más): los
codifican ciertas moléculas de adhesión intercelular.
Para que se produzca la expansión clonal de los linfocitos Tн se necesita
un incremento en la expresión del gen de la interleuquina 2 (IL-2) y de su
receptor (IL-2R). en esta tarea interviene una serie de proteínas reguladoras y
factores de transcripción que se unen a secuencias especificas de la zona 5 no
codificadora (promotor/intensificador) de los correspondientes genes:
·
Complejo
AP1 (c-Fos+c-Jun): se une al elemento TRE.
·
Factor
nuclear NF-AT.
·
Factor
{AP1+NF-AT}, que es específico de las células T: se une al elemento ARRES.
·
Complejo Oct-1+Oct-2+OAP: se une a OBM.
·
Factor
NF-kB: se une a la secuencia kB-RE.
Anergia Clonal
La unión de un linfocito Tн con un complejo péptido-MHC II de una célula
presentadora de antígeno puede conducir a dos tipos de respuestas opuestas:
·
Activación
y expansión clonal
·
Anergia
clonal
La anergia clonal es la incapacidad proliferativa de un linfocito tras un
contacto con el complejo péptido-MHC, y se debe a la carencia de la señal
coestimulatoria proporcionada por interacción entre CD28 del linfocito Tн y B7
de la APC. No se trata de una mera no-repuesta pasiva, sino que la anergia es
un estado activo de no proliferación.
Poblaciones periféricas de
células T maduras
Células T α β
Un 90-95% de las células T periféricas
son de tipo α β (o sea, TCR2), existiendo una proporción de
+CD4+ doble que las CD8+. En general, las CD4+ funcionan como células T
coadyuvantes (Tн) y las CD8+ lo hacen como T citotóxicas (Tc), aunque parece
que ambas poblaciones expresan el mismo repertorio de segmentos variables (Vα y Vβ).
Linfocitos T vírgenes.
Las células T CD4+ y T CD8+ vírgenes
inmunocompetentes que acaban de madurar abandonan el timo y entran en
circulación en un estado de reposo (G0 del ciclo celular). Se caracterizan por
:
·
Bajos
niveles de moléculas de adhesión.
·
Altos
niveles del receptor de alojamiento (homing) llamado L-selectina, que permite
unirse a la dirigina (addressin) vascular de las vénulas de endotelio alto
(HEV) de los ganglios linfáticos. Esto permite la extravasación del linfocito
virgen hasta el interior del ganglio a partir de la circulación.
·
Expresan
la isoforma de alto peso molecular de CD45 (llamada CD45RA), implicada en la
transducción de la señal de activación.
Linfocitos T efectores
Unas 48 horas después de su activación, la célula T convierte en un blasto
y comienza a proliferar en el ganglio linfático, diferenciadose al cabo de 5-7
días en una subpoblacion de células efectoras especializadas y otra
subpoblacion de T de memoria. Las células T efectoras pueden ser de 3 tipos
funcionales diferentes:
·
Tc:
son las T matadoras (citotóxicas), que suelen ser fenotípicamente CD8+.
·
Tн1:
son; las denominadas T inflamatorias, y su papel estriba en activar a
macrófagos. Suelen ser fenotípicamente CD4+.
·
Tн2:
denominadas T colaboradoras o coadyuvantes en sentido estricto, especializadas
en secretar ciertas citoquinas que son esenciales en la activación de celular B
y T. suelen ser CD4+
Linfocitos T de memoria
·
Los
T de memoria surgen como subpoblaciones diferenciadas a partir de la
proliferación de T vírgenes y T efectores durante una respuesta primaria.
·
Permanece
en reposo (fase G0) durante mucho tiempo (hasta 30 años o más), como una
subpoblacion expandida, una vez que ha declinado la subpoblacion ‘’hermana’’ de
células T efectoras.
·
Esta
preparadas para responder de un modo más rápido e intenso cuando se vuelven a
encontrar con el antígeno.
·
En
general poseen el mismo tipo de moléculas de membrana que los T efectores
correspondientes.
·
Al
igual que los T vírgenes recirculan continuamente entre la sangre y la linfa,
pero al carecer de L-selectina y presentar otras moléculas de adhesión, su
patrón de recirculación es distinto: al carecer de L-selectina, no se unen a
las vénulas de endotelio alto (HEV) de los ganglios.
Células T y δ
Estos linfocitos no fueron descubiertos hasta 1986, en
que se reconocieron como una pequeña población de células T periféricas que
expresan CD3 pero no el “típico” receptor TCR α β.
Constituye del 5 al 10% de los T periféricos, y del 1 al
3% de los residentes en ganglios y otros órganos linfoides.
Estos linfocitos epiteliales no recirculan, sino que son
residentes fijos en esos tejidos epiteliales. Lo curioso es que en cada tipo de
epitelio la población residente de T
y δ muestra un repertorio muy limitado de
reordenaciones de segmentos variables; además proceden de “oleadas” distintas
surgidas durante la vida fetal.
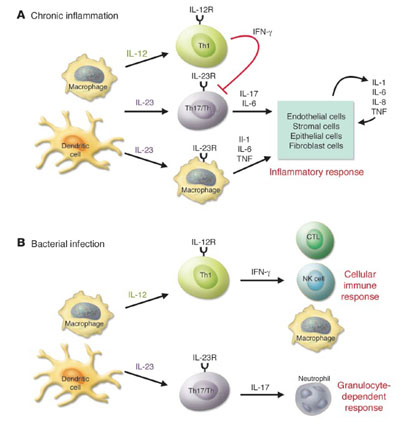
TH17
Los linfocitos Th17 son
distintos a los Th1 y Th2 y se caracterizan por producir IL-17, TNF-alfa, IL-6,
IL-22 y GM-CSF. Inicialmente estos linfocitos fueron detectados en infecciones
por Borrelia burgdorferi,
pero fue su identificación en enfermedades autoinmunes, lo que causó un gran
revuelo e investigación para caracterizar su proveniencia y función.
Las
células dendríticas activadas después de capturar cierto tipo de antígenos por
la vía de receptores tipo Toll (TLR) producen TGF-beta, IL-6 e IL-23 induciendo
la proliferación de Th17. Estas células, a su vez, secretan IL-17A e IL-17F,
promoviendo el reclutamiento de leucocitos polimorfonucleares en infecciones
agudas o heridas. La IL-17 también promueve la producción de GM-CSF, IL-6 y
TNF-alfa, las cuales sustentan a los linfocitos Th17 que participan en las
lesiones crónicas de enfermedades inflamatorias autoinmunes o causadas por
microorganismos. Estas citocinas también activan a células epiteliales,
endoteliales, estromales y fibroblastos, las cuales a su vez producen mas
mediadores como IL-1, IL-6, TNF-alfa, iNOS, metaloproteinasas y qumiocinas que inducen
inflamación.
.jpg)
Recientemente
se ha demostrado que los linfocitos Th17 también participan en la patogénesis
de enfermedades inflamatorias. Blauvelt (2007) propone, después de una
excelente reconstrucción teórica, que la psoriasis debe ser una enfermedad
Th17. De igual manera, los linfocitos Th17 pudiesen ser importantes en otras
enfermedades o desordenes cutáneos caracterizadas por proinflamación e
inmunopatología.
La
investigación sobre los Th17 ha permitido señalar tres vías independientes y
exclusivos de respuestas inflamatorias: IL-12/ IFN-gamma, IL-4/IL-5/IL-13
e IL-23/IL-17. La identificación de la vía involucrada en las distintas formas
clínicas o estadios de una enfermedad permitirá la aplicación de esquemas
terapéuticos mas precisos y certeros.
.jpg)
{kind=link}